Акустическая коммуникация и распознавание акустических сигналов у саранчовых
Большинство
европейских прямокрылых насекомых, а именно
почти все виды кузнечиковых (Tettigonioidea), многие
виды сверчковых (Grylloidea) и саранчовых (Acridoidea),
активно используют акустические сигналы в
процессе поиска полового партнера (Жантиев, 1980;
Попов, 1985; Huber et al., 1989; Bailey, 1991; Gerhardt, Huber, 2002). У
многих прямокрылых звуковая коммуникация служит
единственным известным средством сближения
особей противоположного пола, находящихся на
значительном расстоянии друг от друга (десятки
метров). Саранчовые (Acridoidea), активно использующие
звуковую коммуникацию в половом поведении,
относятся в основном к двум п/семействам: Gomphocerinae
и Oedipodinae. Звуковая коммуникация Gomphocerinae достигает
наибольшей сложности среди всех представителей
Orthoptera в силу особенностей строения звукового
аппарата и разнообразных стратегий поиска
полового партнера (Faber, 1953; Otte, 1970; Helversen, Helversen, 1983,
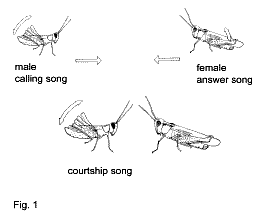
1994). Одиночные самцы спонтанно издают ПРИЗЫВНЫЕ
СИГНАЛЫ (Рис. 1).
Рецептивные самки большинства видов Gomphocerinae
отвечают акустически (ОТВЕТНЫЙ СИГНАЛ) на
призывный сигнал самца. Самец и самка идут друг к
другу, поочередно издавая звуковые сигналы.
Оказавшись рядом с самкой, самец продолжает
издавать звуковой сигнал, причем у одних видов
этот сигнал похож на призывный, тогда как у
других видов СИГНАЛ УХАЖИВАНИЯ в меньшей или
большей степени отличается от призывного
сигнала по временной структуре, интенсивности и
длительности. У некоторых видов сигналы
ухаживания очень сложны и сопровождаются
демонстрационными движениями антенн, головы или
задних ног (видео клипы).
Полевые наблюдения показали, что несколько
самцов могут одновременно ухаживать за одной или
несколькими самками, сидящими рядом. Таким
образом, самка может сравнивать и производить
выбор на основании сигнала ухаживания. Такая
стратегия создает предпосылки для усложнения
временной структуры сигнала в результате
полового отбора. 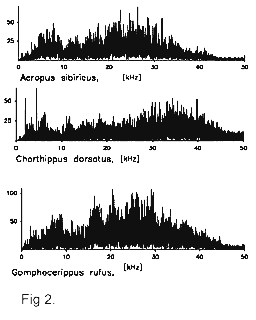
Саранчовые издают
звуковые сигналы посредством трения задних
бедер о надкрылья (фрикционный механизм). На
внутренней стороне заднего бедра располагается
зазубренная жилка (pars stridens), которая трется о
радиальную жилку надкрылья (Рис. 3b, 4). В
результате генерируется относительно
широкополосный звуковой сигнал, частотный
спектр которого может заходить далеко в
ультразвуковую область (Рис. 2). Сигналы разных
видов плохо различаются по огибающей спектра.
Напротив, большие различия наблюдаются по
амплитудно-временным характеристикам
сигнала. 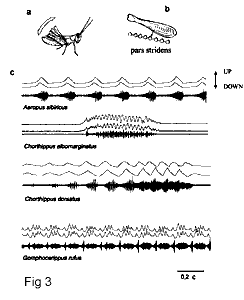
Призывные сигналы Gomphocerinae могут достигать большой сложности по амплитудно-временным параметрам (Бухвалова, Жантиев, 1993; Bukhvalova, Vedenina, 1998; Ragge, Reynolds, 1998; Vedenina, Bukhvalova, 2001; Бухвалова, 2003; Савицкий, 2002, 2004; Vedenina, Helversen, 2009). У большинства исследованных видов ноги движутся c определенным фазовым сдвигом, который может меняться в процессе стридуляции, и, кроме того, сам паттерн движения ног может быть различен (Elsner, 1974). Такое использование двух звуковых аппаратов позволяет саранчовым чрезвычайно усложнять временной паттерн сигнала.
Траектории движений ног регистрировались посредством оригинальной опто-электронной системы, изготовленной в лаборатории N 8 ИППИ РАН на основе описанной в литературе конструкции (Helversen, Elsner, 1977; Hedwig, 2000). Методика одновременной записи звука и движений ног позволяет исследовать связь моторных и акустических характеристик сигнала. Например, если ноги двигаются в противофазе, частота пульсов может удваиваться (Рис. 3, Chorthippus albomarginatus). Если в начале звуковой посылки ноги работают в фазе, а потом начинают двигаться в противофазе, может исчезать расчлененность серий на пульсы, и границы между сериями могут смазываться (Рис. 3, Chorthippus dorsatus, Gomphocerippus rufus). Такое размывание границ между сериями или пульсами часто встречается в сигналах саранчовых. Фазовый сдвиг между ногами у разных особей одного вида может отличаться, в результате чего повышается внутривидовая вариабельность сигналов, которая предоставляет богатый материал для полового отбора (Веденина, 2005).
В сигналах ухаживания
можно выделить больше различных элементов, чем в
призывных сигналах (Helversen, 1986; Vedenina, Helversen, 2003).
Отчасти это может быть связано с особенностями
работы звукового аппарата. В зависимости от
положения бедра разные части зазубренной жилки
могут быть задействованы в генерации звука.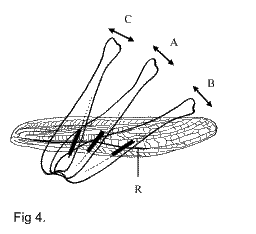
Например, у Ch.
albomarginatus задние бедра, ориентированные почти
горизонтально, колеблются с одной частотой
(генерируя звуковой элемент В, показанный на рис.
5), затем, поднятые на угол около 45о,
колеблются с другой частотой (звуковой элемент А
на рис. 5), и при ориентации почти вертикально
производят третий рисунок движений (звуковой
элемент С на рис. 5) (Рис. 4). Такого сложного
использования фрикционного аппарата не
обнаружено в других группах прямокрылых. В
сигнале Ch. oschei можно выделить, по крайней
мере, пять различных элементов, повторяющихся в
характерном порядке (Рис. 5). Более того, во время
С-элемента самец производит характерный взмах
задними ногами, который возможно служит
дополнительным зрительным сигналом для самки. 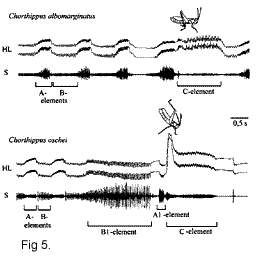
Многочисленные поведенческие и
электрофизиологические исследования,
проведенные за последние три десятилетия,
показали, что для распознавания
конспецифических призывных сигналов саранчовые
используют такие амплитудно-временные параметры
сигнала как длительность звуковых посылок,
частота и длительность пульсов в посылке,
соотношение интенсивности разных пульсов и т.д.
(Helversen, 1972; Helversen, Helversen, 1983; 1987; 1994; Веденина,
Жантиев, 1990; Stumpner, Helversen, 1992, 1994;). В поведенческих
экспериментах самкам саранчовых проигрывали
модельные сигналы с измененными параметрами и
оценивали привлекательность сигнала по
звуковому ответу самки. Более сложные по
временной структуре сигналы ухаживания
практически не исследовались в подобных
экспериментах, так как самки редко издают
ответный сигнал ухаживающему самцу. Реакцию
самки на ухаживание можно оценить на основании
того, допускает она копуляцию или нет. В случае
отказа самка может, тем не менее, оставаться
рядом с ухаживающем самцом, который в конце
концов может достигнуть успеха после нескольких
попыток спариться. 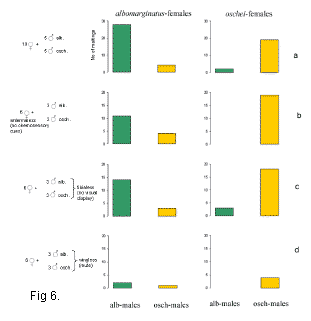
Мы исследовали реакцию самок двух близкородственных видов саранчовых Ch. albomarginatus и Ch. oschei на ухаживание кон- и гетероспецифических самцов. Сигналы ухаживания этих видов хорошо различаются по нескольким амплитудно-временным параметрам звука и демонстрационным сигналам (Рис. 5). Тем не менее, известны случаи гибридизации между этими видами в природе. В лаборатории в условиях свободного выбора самки предпочитали спариваться с конспецифическими самцами, хотя в 10-20 % случаев они копулировали с гетероспецифическими самцами (Рис. 6 а). Кроме того, мы исследовали, какой из компонентов сложного сигнала ухаживания (химический, зрительный или звуковой) играет ключевую роль в распознавании конспецифического сигнала (Рис. 6, b-d). Результаты опытов показали, что наибольшее влияние на реакцию самок оказывает отключение звукового канала связи. Самцы с ампутированными надкрыльями и крыльями производили стридуляционные движения ногами, но не генерировали звук. Они пытались копулировать с самками так же активно, как и интактные самцы. Однако процент копуляций был очень низкий по причине частых отказов самок (Рис. 6 d).
В настоящее время продолжается работа по исследованию информативных элементов песни, которые играют ключевую роль в распознавании конспецифического самца в группе Ch. albomarginatus. Тот факт, что несмотря на большие различия в сигналах ухаживания двух близких видов, самки, тем не менее, иногда совершают ошибки, выбирая гетероспецифических самцов, требует дальнейших исследований.